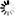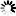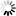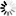Nature全文編譯!揭示新冠病毒刺突蛋白在完整病毒顆粒上的結構和分布
新型冠狀病毒SARS-CoV-2屬于β冠狀病毒屬,是一種包膜病毒,含有較大的由核衣殼蛋白(N)包裹的正義RNA基因組。三個跨膜蛋白被整入病毒脂質包膜:刺突蛋白(S)和兩個較小的蛋白,即膜蛋白(M)和包膜蛋白(E)。當通過低溫電鏡(cryo-EM)成像時,β冠狀病毒呈近似球形顆粒,直徑在100納米上下浮動,內含致密的病毒質(viroplasm),由突脂質雙層包圍著,S蛋白三聚體(下稱S三聚體)從脂質雙層中突出。SARS-CoV-2的S三聚體結合到靶細胞表面上的受體ACE2,并介導隨后的病毒攝取和融合。在這樣做的過程中,S蛋白經歷了顯著的結構重排,從融合前的構象切換到融合后的構象。S蛋白融合前和融合后的整體結構在冠狀病毒中是非常保守的。
在感染過程中,冠狀病毒廣泛地重塑細胞的內部膜結構,產生病毒復制細胞器以便在其中進行病毒復制。S蛋白,連同蛋白M和E,被插入到內質網(ER)的膜中,并被運送到內質網-高爾基體中間區室(ER Golgi intermediate compartment, ERGIC)。封裝的基因組出芽到ERGIC中以形成病毒顆粒,隨后將病毒顆粒運送到質膜并釋放出去。S蛋白是通過先在S1/S2位點隨后在S2'位點進行蛋白酶切割,從而為膜融合做好準備。
包括SARS-CoV-2在內的冠狀病毒的S蛋白融合前結構已通過可溶性的分泌形式的S蛋白異位表達,隨后進行純化和cryo-EM成像,得到了廣泛研究。在S蛋白融合前結構中,受體結合結構域(RBD)位于融合核心(fusion core)上方的一個較寬的S三聚體刺突結構的頂部。在含有三個RBD的S三聚體中,每個RBD被一個顯示出一定流動性的N端結構域(NTD)包圍著。在封閉的融合前結構中,所有三個RBD平鋪在刺突結構表面上,在很大程度上封閉了受體結合位點,而在開放的融合前結構中,一個或多個RBD向上抬起,從而暴露受體結合位點。S三聚體的表面發生廣泛的糖基化,每個S蛋白單體有22個潛在的N-糖基化位點。在結合受體ACE2后,從融合前到融合后的結構轉變讓S蛋白的融合肽和跨膜結構域聚集在一個以三螺旋束為中心的長針狀結構的一端。五個N連接的聚糖沿融合后的S三聚體刺突結構的長度間隔分布。
充分理解S蛋白如何發揮作用,以及它們如何與免疫系統相互作用,需要了解病毒顆粒內S三聚體的結構、構象和分布。在一項新的研究中,來自英國醫學研究理事會分子生物學實驗室和德國海德堡大學的研究人員利用cryo-EM方法研究了S三聚體在病毒顆粒表面上的結構、構象和分布。相關研究結果于2020年8月17日在線發表在Nature期刊上,論文標題為“Structures and distributions of SARS-CoV-2 spike proteins on intact virions”。

圖1.SARS-CoV-2病毒產生特征和圖片,
圖片來自Nature, 2020, doi:10.1038/s41586-020-2665-2。
為了避免與病毒濃縮或純化相關的偽影(artefacts),這些作者想要在不濃縮或純化病毒的情形下對來自感染細胞的上清液的SARS-CoV-2病毒進行成像。利用SARS-CoV-2(病毒分離株Germany/BavPat1/2020)感染VeroE6細胞。在感染48小時后,上清液經澄清后用甲醛固定滅活,并在-80℃下儲存。蛋白印跡法(Western blot)顯示,病毒顆粒上大約45%的S蛋白單體在多精氨酸切割位點上切割成S1和S2(圖1a)。固定的上清液通過驟凍玻璃化,并通過cryo-EM成像。固定可能有助于通過交聯讓一些蛋白構象穩定化,但預計不會產生任何新的構象。正如預期的那樣,考慮到細胞上清液中病毒的濃度(約107個噬菌斑形成單位/ml, PFU/ml),他們發現少量的病毒顆粒分散在網格周圍---這些病毒顆粒通過低溫電子斷層掃描(cryo-electron tomography, cryo-ET)成像(圖1b)。
SARS-CoV-2病毒顆粒是近似球形的,到脂質雙層外邊緣的直徑為91±11納米(n = 179)。它們含有與N蛋白相對應的顆粒狀密度,并綴有S三聚體(圖1b,c)。這些特征與利用cryo-EM成像的其他冠狀病毒的特征基本一致。從這種病毒表面上突出的S三聚體有兩種形態--少數是延長的薄結構,這讓人聯想到融合后的構象,而大多數是更寬的結構,這讓人聯想到融合前的構象。這一觀察結果與近期在預印本服務器上發表的一份論文(bioRxiv, 2020, doi:10.1101/2020.03.02.972927)---該論文顯示了用核酸修飾劑β-丙內酯滅活的純化SARS-CoV-2病毒的cryo-EM圖像,在該圖像中僅在病毒表面上觀察到薄薄的突起---形成了鮮明的對比,但與原位觀察到的病毒組裝相一致。
這些作者還收集了SARS-CoV-2感染CALU-3細胞后產生的病毒顆粒的斷層掃描圖,其中Calu-3細胞是一種人肺癌細胞系,在病毒感染上產生的病毒滴度與VeroE6細胞相當。CALU-3細胞產生的這些病毒顆粒的形態和S三聚體在病毒顆粒表面上的外觀與從VeroE6細胞產生的病毒顆粒所觀察到的相一致。蛋白印跡分析表明,約73%的S蛋白為裂解形式。
SARS-CoV-2病毒顆粒含有24±9個S三聚體。這個值低于之前假設S蛋白等距離分布的估計值,這是因為S蛋白在病毒表面分布不均勻。一小群病毒顆粒僅含有很少的S三聚體,而較大的病毒顆粒則含有更多的S三聚體。這些作者從179個病毒顆粒中鑒定出4104個寬的S三聚體和116個薄的S三聚體,并對它們進行子斷層掃描圖平均化(subtomogram averaging)。平均化的結構,在7.7和22埃的分辨率下,分別非常好地對應于之前公布的純化S三聚體在融合前和融合后形式下的結構(圖2a)。總體而言,約97%的S三聚體是融合前形式,3%是融合后形式。融合前和融合后的S三聚體形式似乎是均勻地分布在病毒顆粒中。

圖2.針對完整病毒顆粒上的SARS-CoV-2 S三聚體的結構分析,
圖片來自Nature, 2020, doi:10.1038/s41586-020-2665-2。
這些作者還收集了SARS-CoV-2感染CALU-3細胞后產生的病毒顆粒的斷層掃描圖,其中Calu-3細胞是一種人肺癌細胞系,在病毒感染上產生的病毒滴度與VeroE6細胞相當。CALU-3細胞產生的這些病毒顆粒的形態和S三聚體在病毒顆粒表面上的外觀與從VeroE6細胞產生的病毒顆粒所觀察到的相一致。蛋白印跡分析表明,約73%的S蛋白為裂解形式。病毒表面上的融合前S三聚體可能主要處于封閉構象,它的開放構象可通過ACE2結合誘導或穩定化,或者它在融合前也可能存在開放構象。當用作免疫原時,開放或封閉的構象可誘導不同范圍的抗體,目前人們正在努力產生穩定在其中一種構象的S蛋白構造體。
為了評估S三聚體是否存在于開放和/或封閉的構象,這些作者對S三聚體中的S蛋白單體的RBD區域進行分類。他們發現三種類型:RBD處于封閉位置的S蛋白;RBD處于開放位置的S蛋白;RBD以封閉位置為主但密度有所減弱的S蛋白,這表明存在更多的可移動構象。
考慮到每個S蛋白單體被分配到的類型,這些作者推導出完全封閉的S三聚體結構,以及其中一個RBD是開放的S三聚體結構,這兩者分別代表了3854個融合前S三聚體中的約31%和約55%(圖2b)。他們還發現了少量S三聚體(3854個融合前S三聚體中的約14%)的兩個RBD處于開放構象(圖2b)。這些觀察結果證實在重組S三聚體中觀察到的RBD的開放也發生在病毒表面上,并且穩定在封閉和開放構象中的人工S蛋白構造體都代表了原位存在的結構。因此,這種受體結合位點在原位隨機暴露,并可與ACE2以及與抗體相互作用。
這些S三聚體并不都是直接從病毒表面突出來的。它們可以向膜傾斜90°,盡管傾斜超過50°就不那么有利。這些作者根據S三聚體相對于膜的方向對它們進行分組,并且分別對每組進行平均化。這些平均化的結構表明膜近端莖區域作為一個鉸鏈有足夠的靈活性,允許向各個方向傾斜(圖2c)。
這些作者構建出SARS-CoV-2病毒顆粒的模型,在該模型中,S三聚體的位置、方向和構象是通過子斷層掃描圖平均化確定的(圖2d)。S三聚體似乎是隨機分布在病毒表面上,它們的位置、方向和構象之間沒有明顯的聚類或關系。在SARS-CoV-2中,每1000 平方納米膜表面大約有一個S三聚體,相比之下,在甲型流感病毒中,每100平方納米膜表面大約有一個S三聚體。在SARS-CoV-2中,S蛋白的稀疏分布以及它們主要處于封閉狀態意味著與大流行性流感病毒相比,受體結合可能較少依賴于親和效應。這與S蛋白和ACE2之間的親和力(在nM范圍內)比血凝素和唾液酸之間的親和力(mM范圍)高相一致。
上清液中低濃度的病毒顆粒使得解析出高分辨率的結構變得困難。因此,這些作者通過蔗糖墊層離心沉淀法對這些病毒顆粒進行濃縮。濃縮的病毒顆粒偏離球形形態,但是它們的整體特征仍保留下來。他們對這些病毒顆粒進行了cryo-ET成像和子斷層掃描圖平均化,觀察到的主要是融合前S三聚體,偶爾有融合后S三聚體。在對融合前S蛋白進行分類后,他們只能識別出處于封閉位置的RBD,以及觀察到RBD密度較弱的S蛋白單體。
從感染細胞的上清液中的病毒顆粒主要顯示融合前S三聚體,這些S三聚體處于封閉或開放的融合前構象。通過蔗糖墊層離心法濃縮的病毒顆粒繼續呈現融合前構象,但不再觀察到開放構象。其他研究已表明,用β-丙內酯,而不是甲醛滅活的病毒顆粒主要處于融合后狀態。從膜中純化的S三聚體僅處于封閉的融合前構象和融合后構象,而其他研究已提示著可溶性S三聚體中的開放RBD是在一系列連續不同的位置中發現的。這些觀察表明這些作者在濃縮之前而不是濃縮之后觀察到的S蛋白的開放融合前構象是脆弱的(盡管文中進行了固定),并可能受到純化過程的影響。
這些作者的數據提示著滅活和純化方法可以改變融合前和融合后形式的比例,以及開放和封閉形式的比例。據推測,病毒表面上大量的融合后S蛋白可能通過屏蔽融合前形式保護這種病毒,或者可能將宿主反應轉向非中和抗體。鑒于在完整的病毒顆粒上觀察到小一部分融合后S三聚體刺突結構,這些作者認為這不太可能是病毒在感染過程中的重要防御機制,但它可能是疫苗接種的一個重要考慮因素。基于滅活病毒顆粒的候選疫苗正在開發中。根據不同的制備方式,這些疫苗可能會向免疫系統呈現不同的S蛋白表位,因此它們誘導中和反應的能力也不同。比如,β-丙內酯經常用于疫苗生產(如用于流感病毒亞單位疫苗),但如果融合后S三聚體誘導非中和反應,那么β-丙內酯可能不是SARS-CoV-2病毒S蛋白疫苗配方制備期間用于病毒滅活的最佳選擇。
接下來,這些作者利用cryo-EM對濃縮的SARS-CoV-2病毒進行二維成像,并對那些從病毒顆粒側面突出的融合前S三聚體進行單顆粒分析,從而產生3.4埃分辨率的融合前S三聚體共識結構。對RBD單體進行集中分類和部分信號減法,可將它們分為兩類。與通過cryo-ET發現樣品中不存在開放構象相一致的是,這些作者觀察到81%的S蛋白單體中RBD處于封閉構象,19%的S蛋白單體中RBD的密度較弱,但主要處于封閉位置。他們將所有三個RBD都處于封閉構象的S三聚體(53%的數據)以及至少一個RBD的密度減弱的S三聚體(47%的數據)的結構分別優化到3.5埃和4.1埃的分辨率(圖3a)。這兩種結構高度相似,僅在一個RBD的密度水平上有所不同。他們利用這種具有三個封閉RBD的結構,建立并完善了S三聚體在病毒表面原位的原子模型。

圖3.利用單顆粒重建方法確定SARS-CoV-2 S三聚體在完整病毒顆粒上的結構
圖片來自Nature, 2020, doi:10.1038/s41586-020-2665-2。
在這些作者建立的結構中,S蛋白表面上的聚糖位置得到了很好的解決:在22個預測的N-糖基化位點中,有17個存在密度(圖3b)。其他5個糖基化位點位于無序的NTD環或莖區域,并沒有在高分辨率下解決。在S三聚體的底部,一個清晰的聚糖環在莖區域上形成一個項圈。相比于已公布的可溶性胞外結構域結構,莖區域的密度延伸了2個螺旋轉彎(helical turn)并在更低的分辨率下進一步延長,直到由于莖區域的靈活性而逐漸消失(圖3c)。SARS-CoV-2的S三聚體分布稀疏,可以高度向膜傾斜。這意味著,在頭部區域基部和莖區域的抗原表位將被抗體接觸到,在那里它們得不到聚糖殼的廣泛保護。
這些作者比較了S三聚體的原位結構和以前使用外源表達的純化蛋白獲得的結構。最近的一項針對溶于去污劑膠團的全長S三聚體的研究確定了在可溶性S蛋白胞外結構域三聚體的大多數結構中沒有觀察到的兩個特征:S蛋白的氨基酸殘基14-26存在明確的密度;S蛋白的氨基酸殘基833和853之間形成折疊環(folded loop)。這個環是在S蛋白胞外結構域的封閉構象結構中折疊形成的,并可能在內體(endosome)的低pH條件下折疊。這些作者僅在氨基酸殘基14-26觀察到較弱的密度,但是他們沒有觀察到833-853區域形成折疊結構。這些作者進行過成像的SARS-CoV-2毒株含有廣泛傳播的D614G替換突變,這種突變破壞了這個氨基酸殘基與K854之間形成的鹽橋(圖3d),并可能減少833-854折疊環的折疊。他們沒有觀察到對應于結合型脂質或其他結合型輔助因子的額外密度。S三聚體可能以罕見的構象存在著,但這不是它們在原位時的一般特征。
總的來說,這些作者確定的結構非常類似于通過雙脯氨酸突變進行穩定化的可溶性三聚體胞外結構域在封閉融合前形式下的結構。這對正在使用的重組純化S三聚體用于研究、診斷和疫苗接種提供了重要的驗證,畢竟它們確實代表了S三聚體的原位結構。通過展示S三聚體在病毒表面上在3.4埃分辨率下的結構(圖3),這些數據讓這些作者樂觀地認為,cryo-EM可以用于研究抗體結合病毒顆粒表面上的S三聚體。這樣的研究可能提供中和抗體如何阻斷病毒感染的新見解,特別是靶向S蛋白的膜近端區域的抗體,從而可以為用于疫苗接種的免疫原設計提供信息。
參考資料:
Zunlong Ke et al. Structures and distributions of SARS-CoV-2 spike proteins on intact virions. Nature, 2020, doi:10.1038/s41586-020-2665-2.
 |
 |
 |
| 官網:m.baichuan365.com | 微信服務號:iseebio | 微博:seebiobiotech |
 |
 |
 |
| 商城:mall.seebio.cn | 微信訂閱號:seebiotech | 泉養堂:www.canmedo.com |
此文關鍵字:新冠病毒刺突蛋白
相關資訊
- 國務院:支持高校和科研院所專業技術人員在職和離崗創業
- 越運動越聰明! Nature:“運動員血液”能讓久坐不動的小鼠滿血復活!
- 德國投入50億歐元以增強科研競爭力
- Science子刊:獨立于膽固醇的遺傳因子SVEP1,或是人類心腦血管代謝疾病的源頭!
- Neurology:太低的LDL膽固醇可能會增加出血性中風的風險
- 如何選擇合適的細胞培養基
- 脫硫凈化劑---環保產品
- 巰基活性聚乙二醇 (Thiol Reactive PEGs) - Nanocs---PEG修飾劑(3)
- 輔助試劑|Auxiliary reagents|Lumiprobe|西寶生物
- 「西寶生物」專業提供美國NU-CHEK【脂肪酸標準品】


新進產品
同類文章排行
- 清除人體內衣原體感染的關鍵鑰匙
- iScience:一種對抗遺傳性神經系統疾病的潛在途徑
- 真的有可以中和所有已知COVID-19毒株的抗體?
- 細胞“廢物”使抗癌的免疫細胞恢復活力!
- 男性注意!這種食品攝入過多會導致結直腸癌
- 無路可退!主食中的農藥殘留,可誘發阿爾茨海默病
- Cell出乎意料新發現:原來是這種“瀕死體驗”讓癌細胞惡化
- 研究發現,冠狀病毒的形狀會影響其傳播
- Science Advances新突破:快速高效生產人體免疫細胞的新方法
- 《Nature Genetics》確定10個克羅恩病相關基因
資訊文章
您的瀏覽歷史